Eriosyce
Na łamach TAXON 2019 pojawił się długo oczekiwany i ważny artykuł, poświęcony badaniom molekularnym nad tym największym rodzajem południowo amerykańskim, ich wynikom i wynikającym z tego zmianom taksonomicznym "Molecular phylogeny of the large South American genus Eriosyce (Notocacteae, Cactaceae): Generic delimitation and proposed changes in infrageneric and species ranks" autorstwa zespołu Pablo C. Guerrero.
Nie chciałbym, aby moje rozważania były przyjęte jako krytyka pracy, bo praca jest świetna i niezwykle potrzebna, ale jako przyczynek do dalszych analiz.
Dotychczas bazowaliśmy na badaniach Nyffeler & Eggli (2010) opartych wyłącznie na jednym markerze trnL-trnF, z którego widać było, że rodzaje Eriosyce, Islaya, Pyrrhocactus i Neoporteria (z Horridocactus, Thelocephala) są blisko spokrewnione.
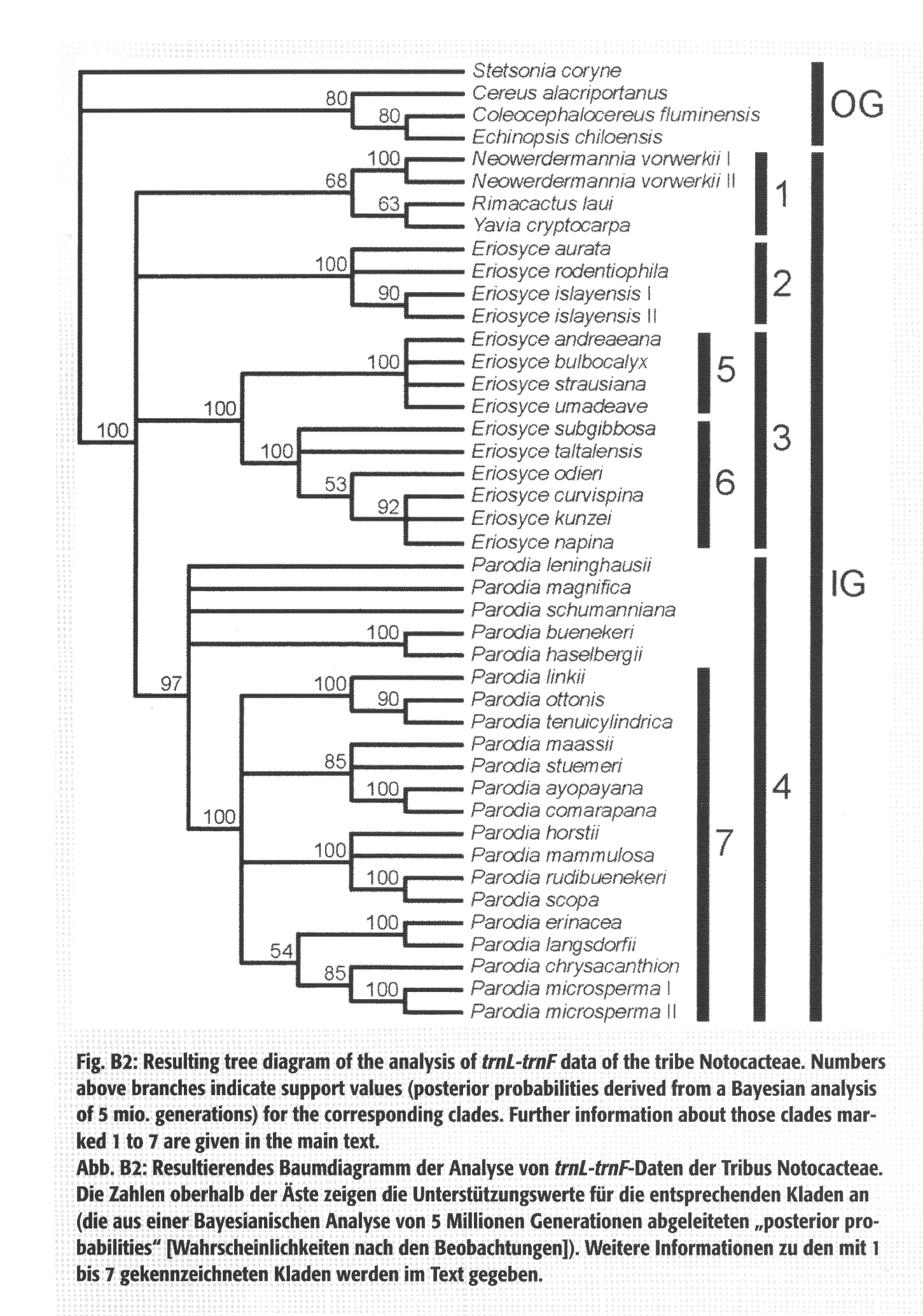
Kladogram Nyffeler & Eggli (2010)
W publikacji zespołu Guerrero dostępne jako załączniki do opracowania są kladogramy wykonane przy użyciu kilku markerów DNA, i to jest bardzo ważne, gdyż zwiększa szanse na prawidłowe rozpoznanie.

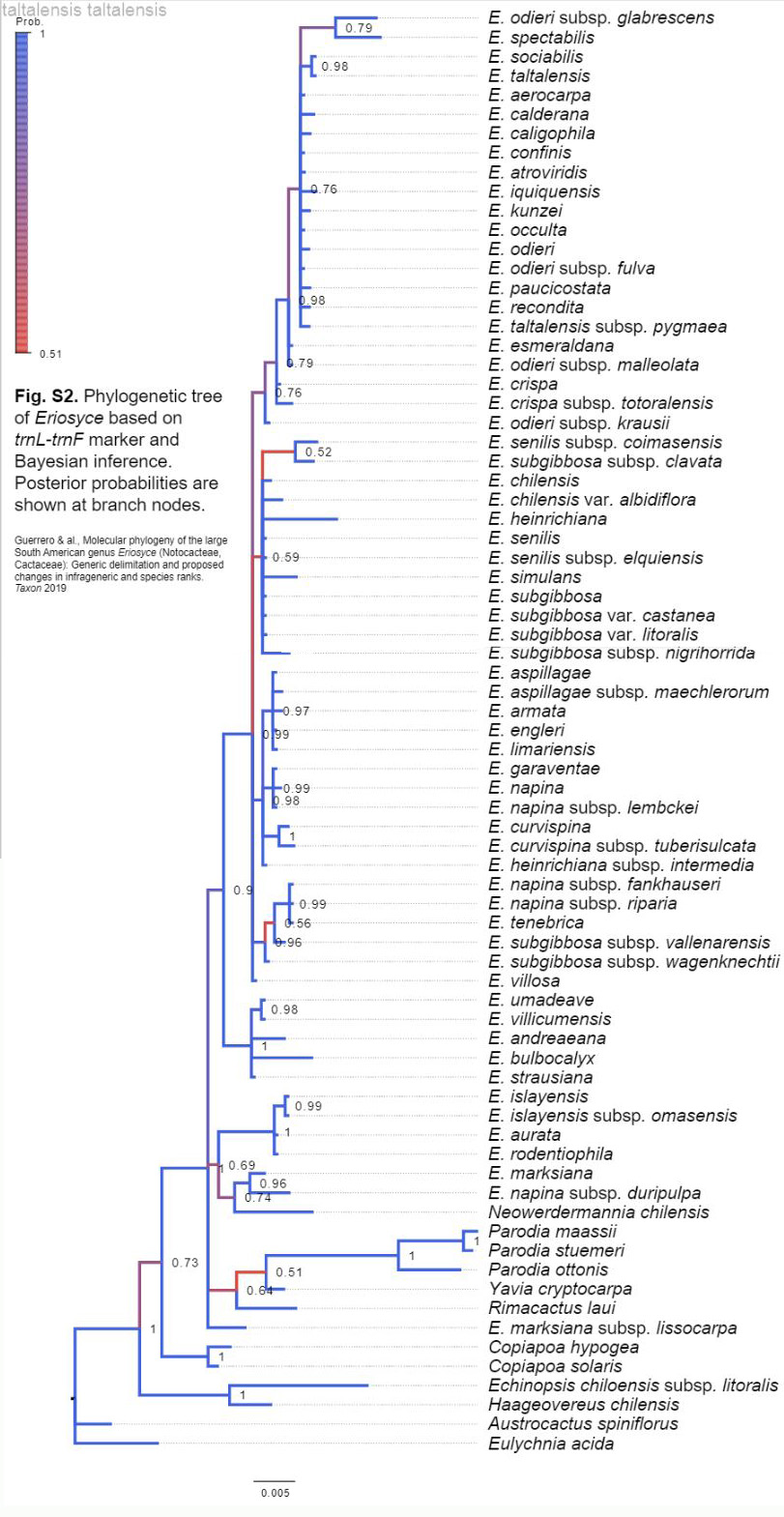
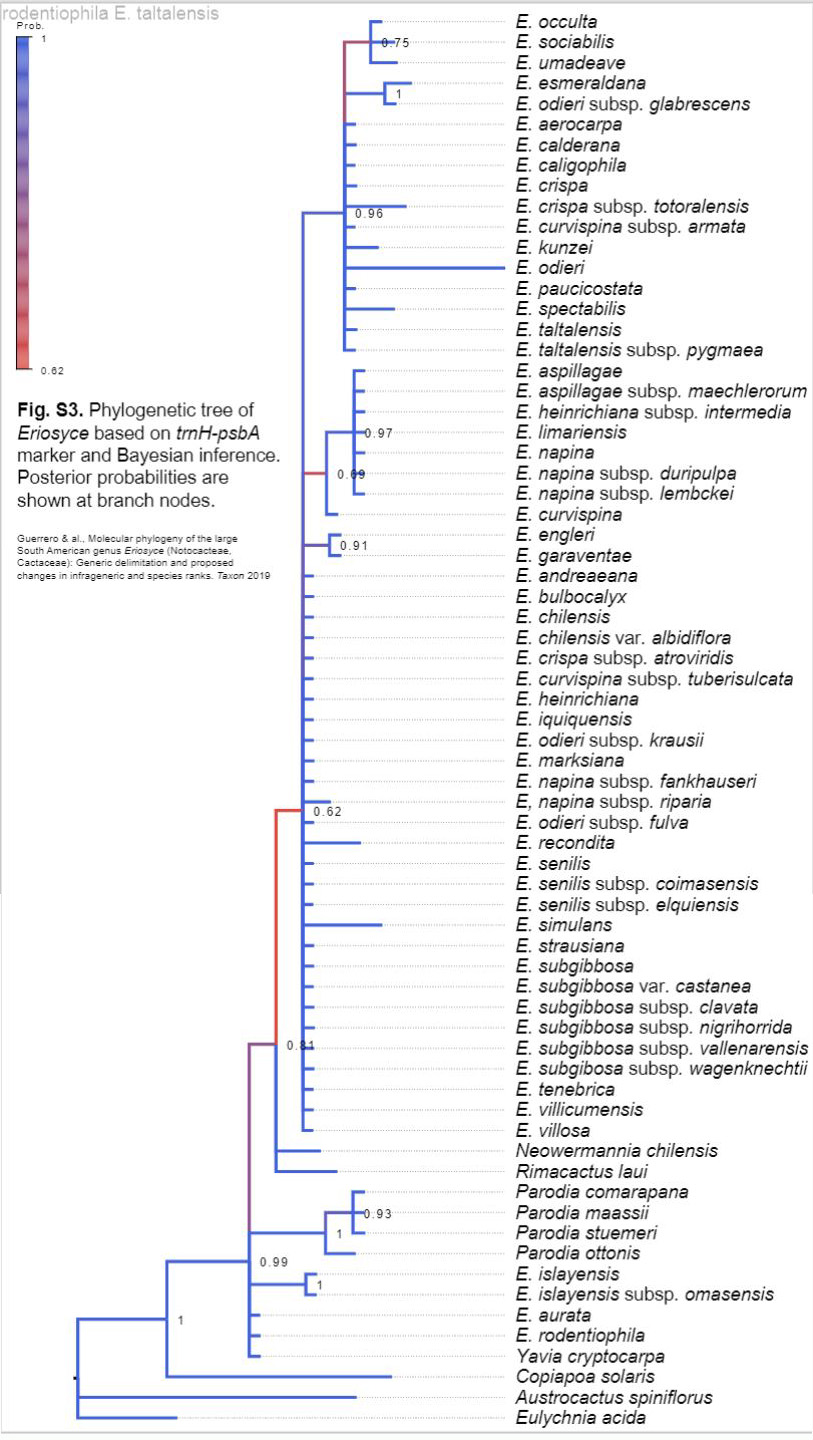



Na podstawie złożenia powyższych kladogramów, otrzymano kladogram wynikowy:
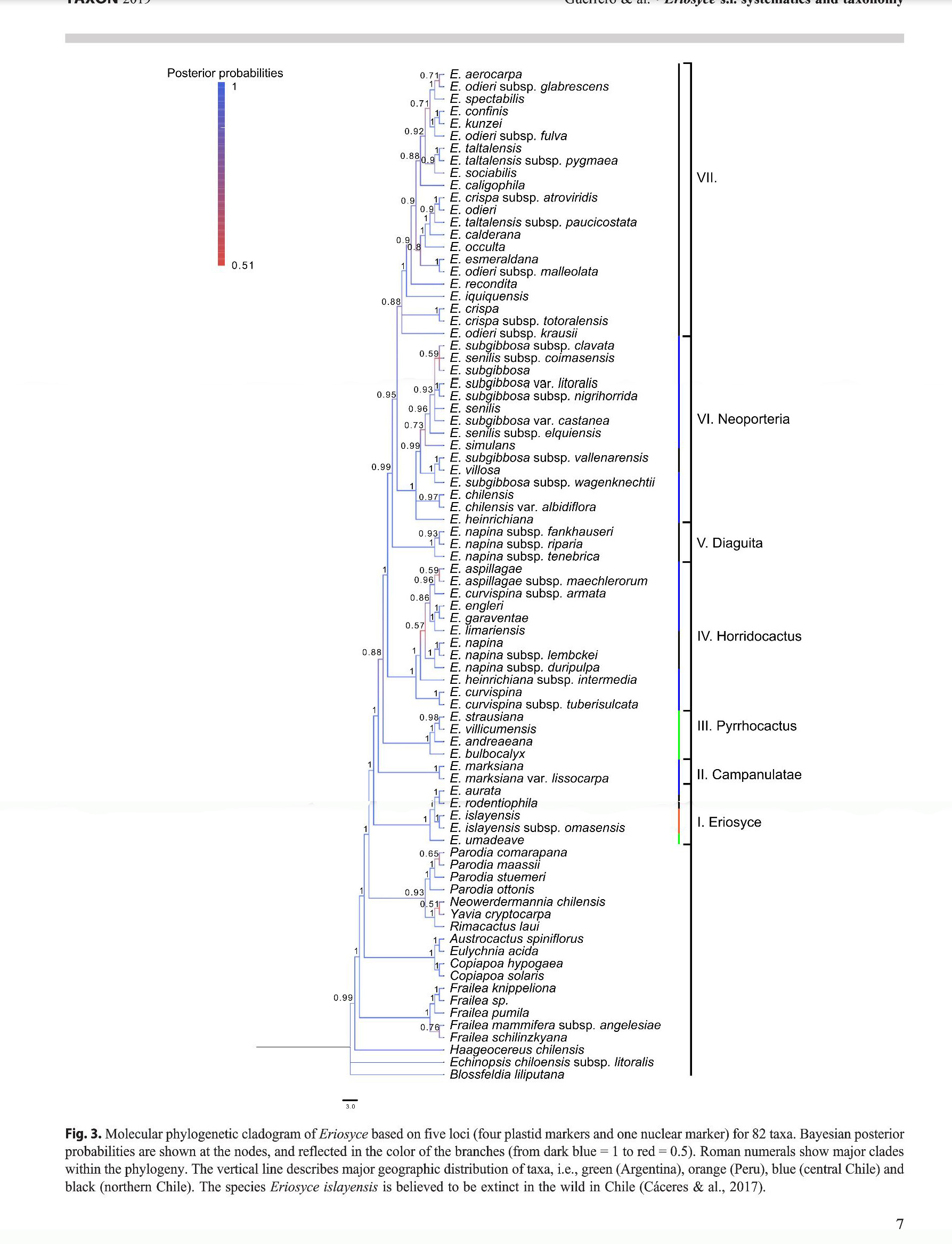
Kladogram wynikowy jest podstawą do dalszych wniosków autorów:
1. utworzono gatunek Eriosyce glabrescens w miejsce Eriosyce odieri subsp. glabrescens
2. potwierdzono, że E. confinis jest synonimem E. kunzei F.Ritter, co sugerowałem w moim artykule The problematic Horridocactus kunzei, przy czym nie został poddany badaniom żaden egzemplarz, wskazywany przeze mnie jako potencjalny Eriosyce kunzei = Echinocactus kunzei.
3. utworzono gatunek E. fulva w miejsce E. odieri subsp. fulva
4. utworzono gatunek E. atroviridis w miejsce E. crispa subsp. atroviridis
5. utworzono gatunek E. malleolata w miejsce E. odieri subsp. malleolata
6. utworzono gatunek E. coimasensis w miejsce E. senilis subsp. coimasensis
7. utworzono gatunek E. litoralis w miejsce E. subgibbosa var. litoralis
8. utworzono gatunek E. nigrihorrida w miejsce E. subgibbosa subsp. nigrihorrida
9. utworzono gatunek E. castanea w miejsce E. subgibbosa var. caastanea
10. utworzono gatunek E. elquiensis w miejsce E. senilis subsp. elquiensis
11. utworzono gatunek E. fankhauseri w miejsce E. napina subsp. fankhauseri
12. utworzono gatunek E. riparia w miejsce E. napina subsp. riparia
13. utworzono gatunek E. tenebrica w miejsce E. napina subsp. tenebrica
14. utworzono gatunek E. armata w miejsce E. curvispina subsp. armata
15. utworzono gatunek E. duripulpa w miejsce E. napina subsp. duripulpa
16. utworzono gatunek E. spinosior w miejsce E. heinrichiana subsp. intermedia
17. uznano, że E. napina subsp. lembckei jest tożsama z E. napina
18. uznano, że synonimem E. malleolata są Thelocephala weisseri A.E.Hoffmann & Helmut Walter, oraz Thelocephala malleolata var. solitaria F.Ritter
19. uznano, że Pyrrhocactus eriosyzoides var. domeykoensis F.Ritter, jest tożsamy z E. atroviridis
20. uznano, że Neoporteria castanea var. tunensis F.Ritter, jest tożsama z E. castanea
21. uznano, że Neoporteria jussieui Britton & Rose, Neoporteria dimorpha (F.Ritter) Donald & G.D.Rowley, Neoporteria setosiflora (F.Ritter) Donald & G.D.Rowley, oraz E. heinrichiana var. setosiflora (F.Ritter) Katt. są tożsame z E. spinosior
22. uznano, że E. napina subsp. challensis jest tożsama z E. duripulpa
Moje uwagi:
Z porównania powyższych kladogramów widać, jak bardzo zasadne jest posługiwanie się kilkoma markerami, co zwiększa dokładność badań. Niemniej istnieje prawdopodobieństwo, że i tak popełniony zostanie jakiś błąd, bo jeszcze inne markery (nie zastososwane w tych analizach) mogłyby wskazać na inne połączenia. Stosowane dotychczas w większości wypadków badania tylko jednego markera trnL-trnF chociaż bardzo pomagają, to jednak nie mogą stanowić jedynej przesłanki uprawniającej do zmian taksonomicznych.
W kladogramie trnH-psbA zastanawiające jest położenie E. umadeave oraz rodzajów Parodia, Neowerdermannia, Rimacactus
W kladogramie trnL-trnF zastanawiające jest ulokowanie E. marksiana subsp. lissocarpa, E. duripulpa i Neowerdermannia chilensis .
Powyższe przykłady pokazują, że wszystkie analizy obarczone są ryzykiem przekłamań, które na pewno były przez autorów analizowane przy układaniu kladogramu wynikowego.
Analiza kladogramów pozwala zauważyć, że poszczególne gatunki (przynajmniej te występujące w Chile) tym bardziej są zbliżone genetycznie, im bliżej siebie rosną, i nie ma to nic wspólnego z wyglądem roślin, np. E. odieri - E. paucicostata, E. confinis - E. fulva
Zrównanie E. napina subsp. challensis z E. duripulpa jest moim zdaniem przedwczesne, bo tak bardzo różnią się wyglądem, że zachowanie podgatunku powinno nastąpić. Zwracam uwagę, że E. napina subsp. challensis nie brała udziału w badaniach DNA, czyli autorzy opracowania bez badań synonimizują takson według własnego zdania.
Zawsze uważałem, że E. fankhauseri, E. riparia, E. tenebrica są sobie bliskie, ale też że nie są powiązane genetycznie z odległą i oddzieloną wysokimi górami E. napina, czyli nie są jej podgatunkami lub też odmianami.
Nie jest mi znana przyczyna, dla której autorzy ograniczyli się ze zmianami w klasyfikacji do części taksonów, pozostawiając inne bez zmian. Dla porównania zamieszczam kladogram wynikowy uzupełniony o nowe nazwy (zaznaczone kolorem niebieskim). Przewiduję, że w niedalekiej przyszłości również zostaną zmienione nazwy innych (zaznaczyłem kolorem zielonym).
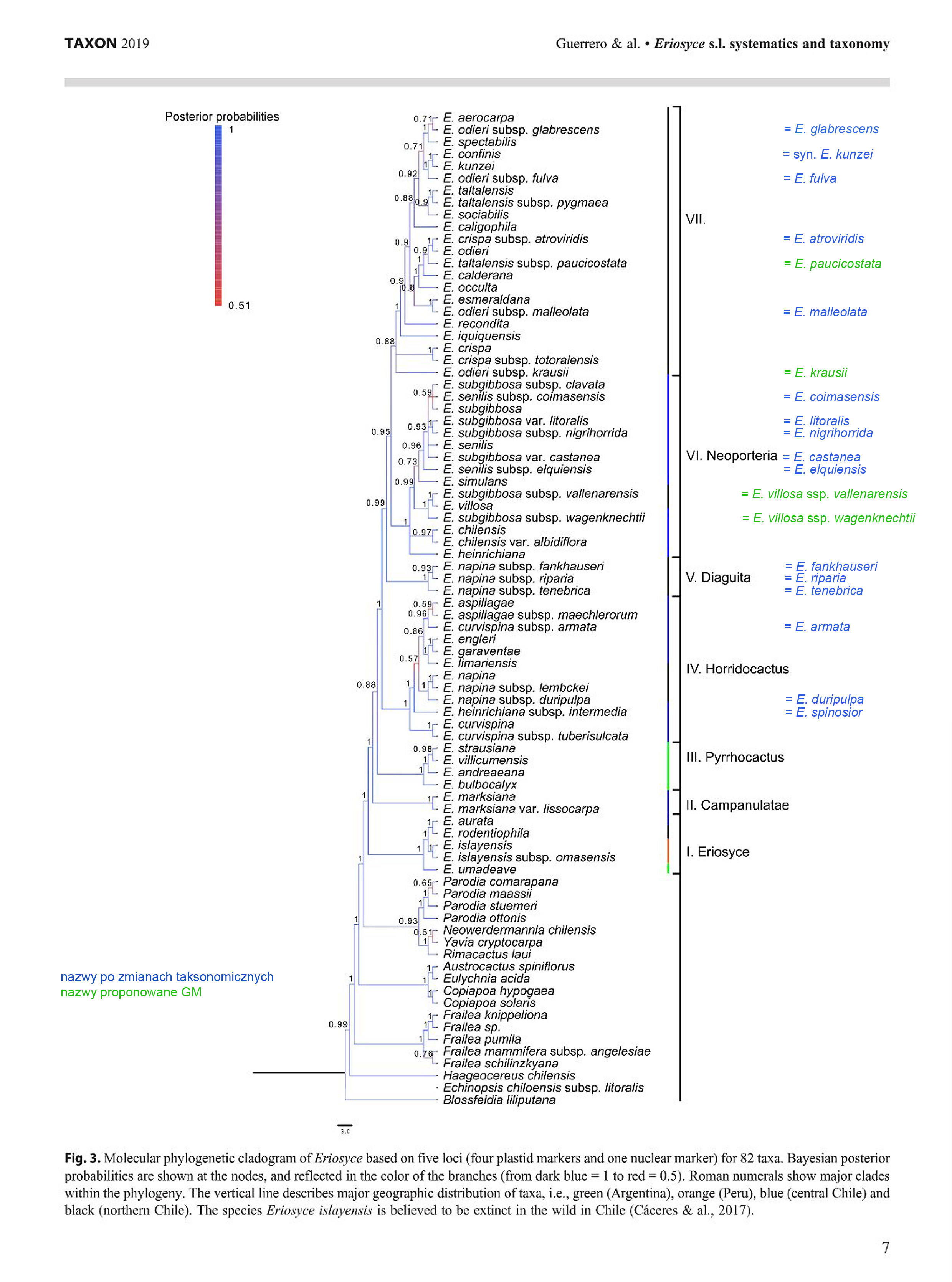
Na koniec jeszcze jedna uwaga.
Analizując drabinkę kladogramu i mając na uwadze wcześniejsze kladogramy, można odnieść wrażenie, że rodzaj Eriosyce nie powinien obejmować wszystkich sekcji w górę, lecz powinien zamknąć się wyłącznie na sekcji I. Sekcję III powinien objąć rodzaj Pyrrhocactus, zaś sekcje IV, V, VI i nienazwana VII, jako stosunkowo spójne, powinny wejść w skład rodzaju Neoporteria, ewentualnie jeszcze bardziej rozdzielić, nadając każdej grupie przypisaną do niej nazwę jako rodzajową z dopisaniem dla nienazwanej jakąś nazwę. Przecież jeśli spojrzymy na drabinkę kladogramu poniżej sekcji I, to równie dobrze można byłoby włączyć rodzaj Eriosyce do rodzaju Parodia i tak dalej. Wydaje mi się, że autorzy po prostu chcieli uhonorować F.Kattermann`a pozostawiając tak obszerny rodzaj.
Tylko od botaników zależy, czy tak wiele gatunków obejmujący rodzaj Eriosyce zostanie utrzymany, czy też będzie podzielony na mniejsze rodzaje, ale bardziej odzwierciedlające zespoły cech wspólnych. Ja osobiście wolę to drugie rozwiązanie.
Pomimo, że badania były niezbędne i dobrze, że zostały wykonane, to wielka szkoda, że nie zostały objęte badaniem dalsze gatunki. I tak dobrze byłoby porównać z Eriosyce kunzei (confinis) Ritter populacje Eriosyce eriosyzoides z okolic Balala, Pyrrhocactus transitensis z Transito, Eriosyce sp. z okolic Hurtado, Fundina i Tres Cruces (Rivadavia) Horridocactus carrizalensis i Neoporteria huascensis , ponieważ moim zdaniem Eriosyce kunzei Ritter nie jest tożsame z Eriocactus kunzei Foerst., ale z Eriosyce atroviridis - dla mnie jest to ewidentny bąd taksonomów. Zabrakło mi też badań grupy Eriosyce paucicostata z Horridocactus echinus, Horridocactus floccosus i Pyrrhocactus neohankeanus z Quebrada San Ramon. Brak również badań Neoporteria pilispina, Neoporteria aricensis, Neoporteria clavata var. procera, Thelocephala longirapa. .
Zwracam również uwagę, na możliwość powstawania błędów w badaniach molekularnych z jeszcze jednego powodu. Otóż wszystkie próbki materiałów genetycznych pobierane były z ogrodów botanicznych oraz dużych kolekcji, gdzie rośliny są, dla zachowania gatunku, mnożone generatywnie. Znamy doskonale fakt, że gatunki chilijskie bardzo łatwo się krzyżują, a obecności zapylaczy w szklarniach trudno się pozbyć. Zatem mnożone od dawna rośliny mogą posiadać część genów obcych dla swojego gatunku, a to w oczywisty sposób prowadzi do fałszywych wyników badań. Uważam, że do badań molekularnych powinno się pobierać wyłącznie rośliny z naturalnego środowiska, bądź rozmnażane z nasion zebranych w naturze.
Problem Neowerdermannia chilensis
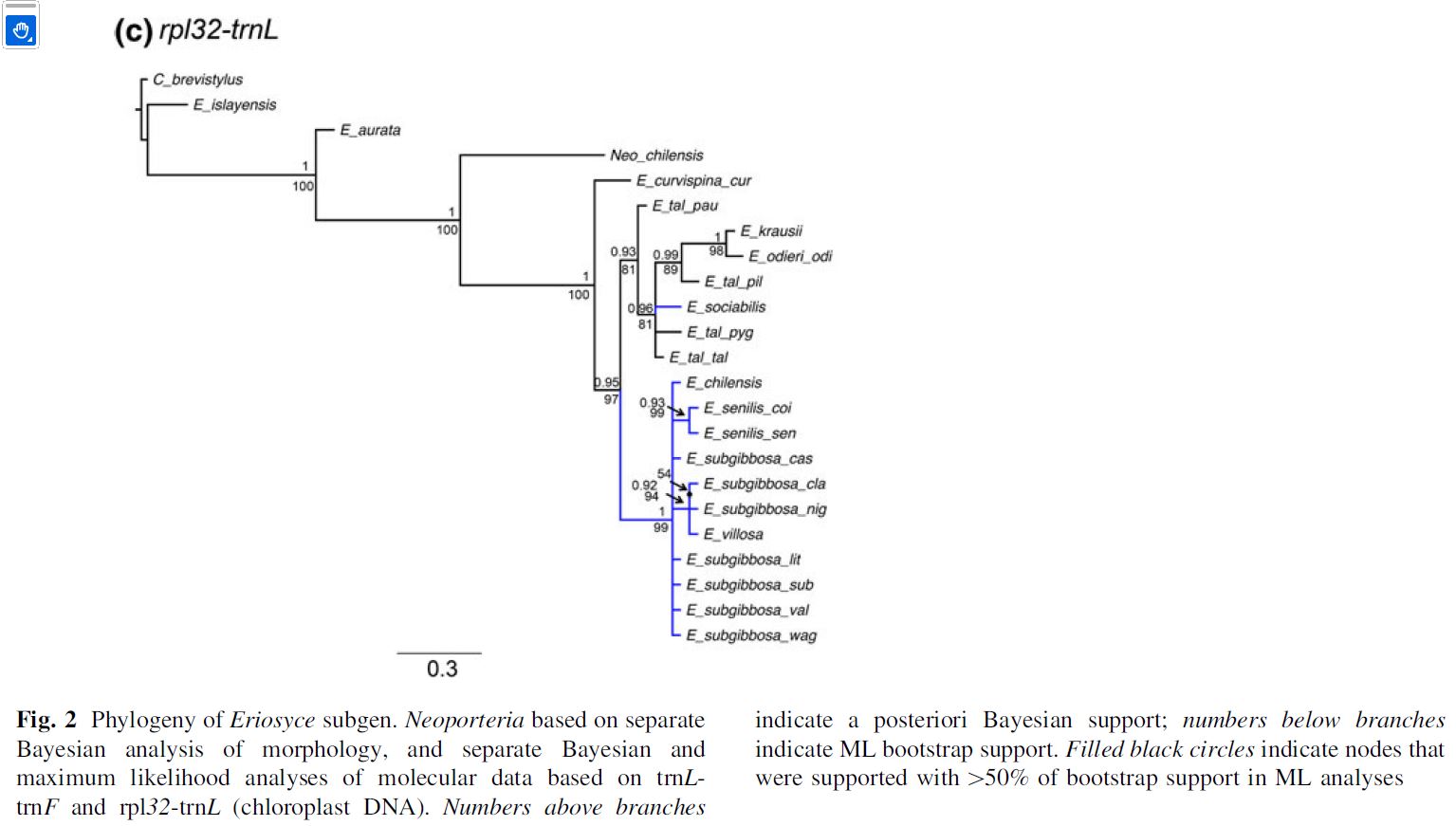
We wcześniejszych badaniach P.Guerrero (2011) - https://www.researchgate.net/publication/230760862
Neowerdermannia chilensis została umieszczona w kladogramie między Erioosyce aurata a Eriosyce curvispina. W przedstawionych wyżej badaniach trnL-trnF Fig. S2 N. chilensis jest w tej samej grupie co E. aurata i E. marksiana. W drzewie Fig. S3 opartej o marker trnH-psbA Neowerdermannia chilensis jest razem z Rimacactus laui, na zewnątrz grupy Eriosyce, ale między Eriosyce i Parodia.
W kladogramie Fig. S4 opartym o marker ycf1 również Neowerdermannia chilensis
jest między Eriosyce i Parodia
. W drzewie Fig. S6 jest już Neowerdermannia chilensis
w grupie Parodia. W wynikowym drzewie Fig. 3 Neowerdermannia chilensis umieszczono razem z Parodia
(poza Eriosyce).
Moim zdaniem uczyniono to
celowo, gdyż wcześniejsze położenie Neowerdermannia chilensis
rozbijało (ze względu na cechy morfologiczne) w miarę spójny układ drzewa
Eriosyce. W konsekwencji trzeba by rozbić rodzaj Eriosyce
sensu lato na poszczególne rodzaje (Pyrrhocactus, Eriosyce,
Horridocactus, Neoporteria i.t.d.
Wydawałoby się, że identyczne badania przy użyciu
tego samego markera powinny przynosić identyczne wyniki, bo jeśli nie to takie badania nie miałyby
sensu. Tymczasem w badaniach prowadzonych przez tego samego naukowca (P.Guerrero) z użyciem identycznego
markera rpl32-trnL Neowerdermannia chilensis jest umieszczona w
różnych miejscach w zależności od roku badań. W 2011 badana była grupa Neoporteria, więc położenie Neowerdermannia chilensis nie miało wpływu na wynik, podczas gdy już w następnym badaniu dotyczącym całej grupy Eriosyce pierwszy wynik powodował że rodzaj Eriosyce stawałby się polifyletyczny. Moim zdaniem poprawiono "ręcznie" położenie kłopotliwego taksonu dla udowodnienia swojej tezy.
Zgodnie z ostatnim opracowaniem P. Guerrero & H. Walter z 2022 (Phytotaxa 550 (2): 079–098) niektóre taksony zostały przeniesione z wcześniejszych rodzajów/podrodzajów do innych, np. Eriosyce heinrichianus z grupy Horridocactus do Neoporteria, czy Eriosyce sociabilis z grupy Neoporteria do jeszcze nienazwanej VII grupy. Pozostawienie wielkiego worka Eriosyce sensu lato pozwala na pozbycie się kłopotu z taksonomią.